Содержание:
Советский Энциклопедический Словарь. 1980
Популярная медицинская энциклопедия. 1981
Большая российская энциклопедия. 2017
Энциклопедический Словарь. Том 2. — Москва, 1953—1955. — Стр. 467
НАСЛЕДСТВЕННОСТЬ. По К. А. Тимирязеву, Н.— сохранение и передача сходного как во внешнем или внутреннем строении, так и в химико-физических особенностях и в жизненных отправлениях организмов. По Т. Лысенко, Н.— свойство организма требовать определённых условий для своей жизни, своего развития и определённо реагировать на те или иные условия; Н. является как бы концентратом воздействий условий внешней среды, ассимилированных организмами в ряде предшествующих поколений.
 Советский Энциклопедический Словарь. — Москва, 1980. — Стр. 874
Советский Энциклопедический Словарь. — Москва, 1980. — Стр. 874
НАСЛЕДСТВЕННОСТЬ, свойство организмов повторять в ряду поколений сходные типы обмена веществ и индивидуального развития в целом. Обеспечивается самовоспроизведением материальных единиц Н.— генов, локализованных в спецефич. структурах ядра клетки — хромосомах. Вместе с изменчивостью Н. обеспечивает постоянство и многообразие форм жизни и лежит в основе эволюции живой природы.
Популярная медицинская энциклопедия. — Москва, 1981. — Стр. 394—398
НАСЛЕДСТВЕННОСТЬ — присущее всем организмам свойство передавать потомству характерные черты строения, индивидуального развития, обмена веществ, а следовательно, состоянии здоровья и предрасположенности ко многим заболеваниям. По наследству могут быть переданы признаки не только нормального, но и измененного болезненного, патологич. состояния организма. Здоровье человека, как и предрасположенность к заболеваниям, в значительной мере наследственно обусловлены. Являясь важнейшим общебиологич. свойством живого, Н. обеспечивает многообразие форм живых существ. В то же время изменение конкретных наследственных свойств, закономерно происходящее за счет процесса изменчивости, сопряженное с процессом естественного отбора лучших форм (см. Эволюционное учение), обеспечивает непрерывность процесса эволюции живых существ на Земле.
Н. возможна благодаря проявлению специфич. черт устройства генетического аппарата, реализации его в процессе индивидуального развития (см. Онтогенез) в определенные признаки и свойства морфологич., физиологич. или биохимич. организации живых существ, а также передачи точного строения генетич. аппарата потомкам. Выяснение закономерностей наследования — важнейшее достижение материалистич. науки.
Основные понятия и законы учения о наследственности. Одним из основополагающих принципов учения о Н. является характеристика наследственных задатков — генов как дискретных (отдельных) частиц живой материи, к-рые определяют признаки и свойства организма в процессе его развития. Организм получает наследственные задатки — гены — от родителей в результате полового процесса — скрещивания или деления клеток исходного организма при бесполом размножении (см. Paзмножение). В клетках тела взрослого организма каждый ген имеет пару (аллель). При созревании половых клеток (гамет) аллели гена расходятся в разные гаметы. Т. о., каждая гамета несет один ген из пары. Эта закономерность, известная под названием закона чистоты гамет, была вскрыта Г. Менделем. При оплодотворении половые клетки отца (сперматозоиды) и матери (яйцеклетки) сливаются, образуя новую клетку — зиготу, в к-рой для каждого из признаков имеется уже пара задатков (генов) — один отцовский, другой материнский. Т. о. в будущем новом организме наследственные признаки обусловлены парой генов, полученной от обоих родителей. Вместе с тем в организме ребенка признаки каждого из родителей проявляются неодинаково. Так, напр., известно, что внешний облик, детали обмена веществ, черты характера ребенка могут в большей степени соответствовать признакам одного из родителей. Это, как установил Г. Мендель, связано с тем, что существует два типа наследственных задатков (генов) — сильные (доминантные) и слабые (рецессивные). Признаки, определяемые доминантными генами, обязательно проявляются в процессе индивидуального развития организма, действие рецессивных генов в процессе их взаимодействия с доминантными подавляется. По предложению Г. Менделя, доминантные факторы обозначают прописными буквами (А, В, С и т. д.), а рецессивные — строчными (а, Ь, с и т. д.).
Поскольку доминантный фактор А подавляет в зиготе действие рецессивного фактора а, то из этой зиготы (Г. Мендель назвал эти клетки гетерозиготными) разовьются организмы, внешний вид к-рых будет определяться лишь фактором А. Только в организмах, клетки к-рых содержат по паре рецессивных факторов а, определяемый признак имеет внешний вид (фенотип), определяемый рецессивными факторами (генами). Организмы, в клетках к-рых парные гены одинаковы (оба доминантные или оба рецессивные), называют гомозиготными. Если бы родители различались только по одному признаку, нетрудно представить схему сочетания признаков (рис. 1).
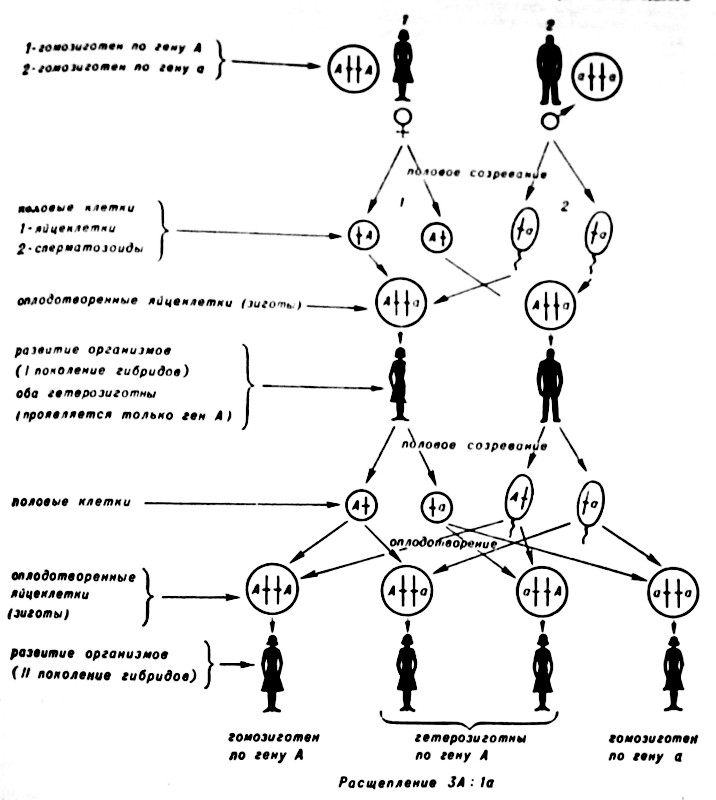
Рис. 1. Цитологические основы моногибридного скрещивания. В неполовых клетках материнского организма (♀) ген, по которому различают генотипы родителей, представлен в каждой из гомологичных хромосом доминантной формой (A), в отцовском организме (♂) рецессивной (а). Если хромосомы в клетках содержат пару одинаковых генов, то их называют гомозиготными по этим генам. Таким образом, материнский организм (1), указанный на данном рисунке, гомозиготен по доминантному гену А, а отцовский (2) — гомозиготен по рецессивному гену а. При половом созревании образуются клетки (яйцеклетки у материнского организма и сперматозоиды у отцовского), которые содержат по одной копии каждой xpoмосомы, и во время оплодотворения происходит восстановление парного (двойного) набора хромосом. Важно, что при этим все яйцеклетки (1) несут по одному гену А, точно также все сперматозоиды (2) имеют гены а. При оплодотворении яйцеклетки оплодотворяются сперматозоидами, в результате чего получается клетки (зиготы), вновь несущие двойной набор хромосом с доминантным и рецессивным геном (Aa) — гетерозиготные клетки. При развитии организмов, содержащих гетерозиготные клетки, внешне проявится только признак, контролируемый доминантным геном (А). Половые клетки гетерозиготных opганизмов (I поколение гибридов) вновь содержат по одной хромосоме, при этом как с доминантным, так и с рецессивным генами образуют яйцеклетки, и сперматозоиды. При скрещивании этих организмов имеется вероятность образования не одного типа зигот, а четырех. При этом рецессивный ген внешне проявится только у особей, несущих в своих клетках пару рецессивных генов. Во всех остальных случаях внешне проявится только доминантный ген. Особей с проявлением доминантного гена будет в три раза больше, чем с проявлением рецессивного гена. Следует подчеркнуть два обстоятельства, на которые обращал внимание уже Мендель при открытии им закона расщепления. Первое — вероятностный характер этих численных соотношений. Соотношение 3:1 (как и любое другое численное соотношение при скрещивании особей, различающихся уже не по одному гену, как в приведенном примере, а по два, три и большему числу генов) можно получить, если изучить достаточно большое число потомков от скрещивания. И второе — то, что расщепление наступает только во II поколении гибридов. В I поколении все особи (несмотря на их гетерозиготность) имеют одинаковый внешний вид (фенотип).
Явление доминантности широко распространено в природе (оно проявляется и в отношении наследования многих признаков у человека), однако проявление доминантности различно. В ряде случаев имеет место неполное доминирование: в фенотипе потомков частично проявляется признак как одного, так и другого родителя. Хорошо известно явление так наз. множественного аллелизма, при к-ром ряд генов последовательно доминирует один над другим. Признаки, наследование к-рых подчиняется перечисленным закономерностям, принято называть менделирующими (по имени Г. Менделя).
У человека менделирующими признаками являются, напр., альбинизм (отсутствие пигментации, вызываемое рецессивным геном; встречается у всех человеческих рас с частотой 1 на 20—30 тыс. новорожденных), цвет глаз, характер волос (курчавые или гладкие), групповые отличия по различным факторам в крови (см. Группы крови) и др. Законам Менделя подчиняются и гены, обусловливающие наследственные болезни человека.
Изучение закономерностей проявления рецессивных генов у гетерозиготных организмов (Аа) имеет огромное теоретич. и практич. значение. Напр., если рецессивный ген (а) определяет наследственное заболевание, то важно знать, что гетерозиготные организмы (Аа) являются носителями таких скрытых патологич. генов, проявление к-рых в фенотипе их гомозиготных потомков (аа) влечет за собой выявление болезни. В ряде случаев рецессивный ген частично проявляет себя и в гетерозиготном организме. У человека и животных таким образом наследуется альбинизм. Если гомозиготные организмы (аа)— полные альбиносы (отсутствует пигментация кожи, волос и радужки глаз, зрение ослаблено), то гетерозиготные (Аа)— частичные альбиносы (светлая окраска кожи, волос, почти бесцветные глаза и др.).
Экспериментально изучив результаты скрещивания организмов, различающихся по одному, двум, трем и т. д. числу факторов (генов), Г. Мендель сформулировал правила наследования — возможные соотношения проявлений родительских признаков у их потомства. Г. Мендель не знал, что собой представляют наследственные факторы (гены) и где они расположены в клетках, поэтому он считал, что наследственные факторы передаются независимо друг от друга. Позднее было установлено, что гены сосредоточены в ядре клетки и располагаются по длине особых структур, , названных хромосомами. Амер. ученый Т. Морган и его ученики не только доказали связь генов с хромосомами, но и разработали метод изучения взаиморасположения генов в хромосоме. При этом в разных хромосомах может находиться разное число генов. Гены, расположенные в одной хромосоме, передаются при скрещивания сцепленно, совместно (если только не произойдет обмен участками между разными хромосомами). Выяснив, что хромосомы могут обмениваться друг с другом участками за счет перекреста — кроссинговера (см. Ген), Т. Морган установил, что чем дальше друг от друга находятся в хромосомах два гена, тем чаще между ними может произойти кроссинговер (рис. 2).
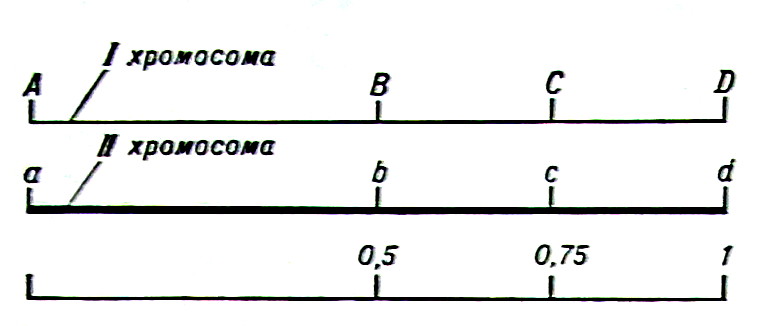
Рис. 2. Генетическая карта гомологичных хромосом, содержащих четыре гена, представленных в доминантных (A, В, С и D) и рецессивных (а, Ь, с и d) формах. Если принять расстояние между генами А и D на генетической карте первой хромосомы за единицу (соответственно этому расстояние а — d на второй хромосоме также составит единицу). то расстояния А — В. В — D, а — Ь и Ь — d будут равны 0,5; а В — С. С — D, Ь — с и с — d — 0.25. Вероятность перекреста в участке между генами А — В и В — D будет вдвое меньше. чем между генами А — D, а между генами В и С eще в два раза меньше, пропорционально тому, во сколько раз меньше длина каждого из участков по сравнению с длиной всего отрезка А — D. Таким образом, изучая частоту перекреста между двумя избранными генами, можно определить относительное расстояние между ними. Это расстояние обычно выражают в долях от всей генетической карты данной хромосомы, длину которой принимают за единицу. Цифры внизу рисунка обозначают относительную частоту перекреста между генами и соответствуют относительному расстоянию между этими генами в хромосомах.
Напр., если расстояние между генами А и D в первой хромосоме и a и d во второй хромосоме в 2 раза больше расстояния между генами A и B, то кроссинговер между этими хромосомами я участке A—D может произойти в 2 раза чаще, чем в участке A—B, а кроссинговер в участках A—B и B—D — в 2 раза чаще, чем в участке B—C или C—D. На этом основании Т. Морган предложил использовать частоту кроссинговера как меру расстояния между генами.
Установлено, что в клетках каждого вида организмов содержится строго определенное число хромосом и, более того, форма и строение каждой отдельной хромосомы строго постоянны. Набор хромосом клеток одного вида (кариотип) имеет одинаковый вид, этот групповой портрет настолько определенен, что опытный специалист по виду набора хромосом может отличить клетки одного вида от другого.
С хромосомами связано еще одно важное свойство живых клеток. Т. Морган установил, что в кариотипе каждой клетки тела есть пара хромосом, определяющих пол особи,— половые хромосомы. По форме половые хромосомы делятся на 2 группы: первые — большой длины и налочковидной формы — были названы Х-хромосомами, а вторые — небольшие, нередко загнутые крючком — Y-хромосомами. У особей женского пола животных, растений и многих насекомых в ядрах клеток тела содержится по две Х-хромосомы, у мужских особей — одна Х- и одна Y-хромосома. Открытие этого правила позволило понять, почему число особей каждого пола примерно равно. Если изобразить процесс образования половых клеток и последующего их оплодотворения, то можно увидеть, что равное число особой обоего пола всегда будет сохраняться (рис. 3).
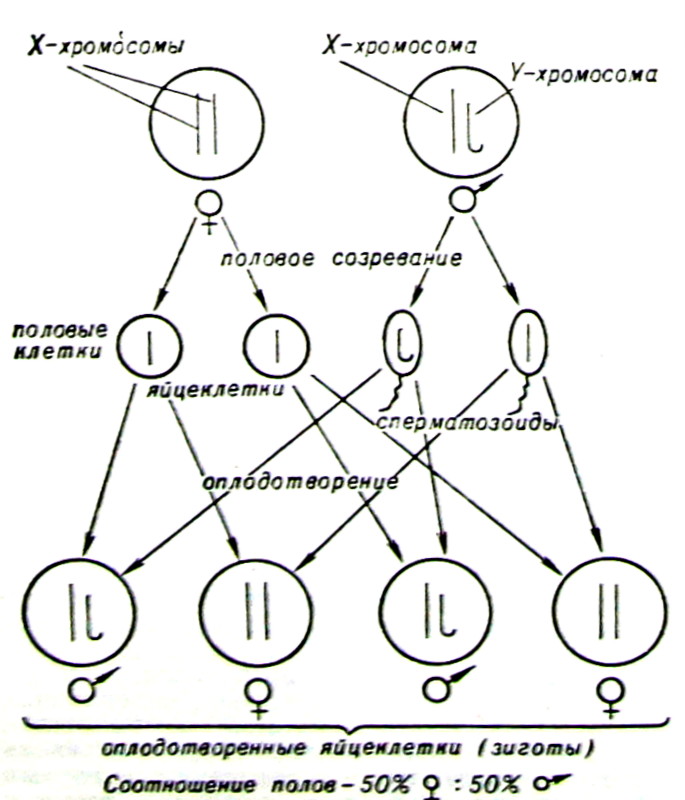
Рис. 3. Хромосомные основы соотношения полов. Клетки самок (♀) обычно содержат пару X-хромосом, а самцов ()— одну Х-хромосому и одну Y-хромосому. Поэтому при созревании яйцеклетки несут по одной Х-хромосоме, а сперматозоиды как Х-хромосому, так и Y-хромосому. При оплодотворении это сочетание приводит к появлению одинакового числа самцов и самок, что повторяется из поколения в поколение.
Всякое изменение числа хромосом в клетках оказывает глубокое влияние на свойства организма. Известны случаи. когда в клетках оказывается на одну или несколько хромосом больше, чем в норме, или же на одну или несколько хромосом меньше. Такие организмы называют анеуплоидными. У человека нехватка или избыток отдельных хромосом приводит к тяжелым наследственным болезням. Кроме того, нередко у многих организмов происходит удвоение, утроение и т. д. всего набора хромосом — кариотипа. Это явление названо полиплоидией. Полиплоидизация у растений, по-видимому, играла большую роль в эволюции растительного мира.
Особенно важными для обоснования теории Н. оказались случаи нарушений в структуре половых хромосом. Возможность точного математич. предсказания признаков аномальных форм стала лучшим способом проверки правильности хромосомной теории Н., при к-ром отклонения от нормы получили законченное объяснение. Одновременно с этим детальному изучению подверглись случаи наследования генов, располагающихся не в половых, а в остальных хромосомах, так наз. аутосомах. Разработан специфич. метод изучения хромосом, к-рый позволяет выявлять индивидуальные черты строения отдельных хромосом на основе использования метода их дифференциальной окраски. С этим методом связывают надежды на выявление сложных случаев обмена участками между хромосомами — так наз. транслокации (см. Мутагенез), что важно для мед. и с.-х. генетики.
Формирование отдельных признаков, как правило, происходит в результате взаимодействия большого числа генов.
В процессе развития организма при формировании его признаков происходит сложное взаимодействие продуктов различных генов. Действие одних генов подавляется других, или признак проявляется только при совместном участии нескольких генов (комплементарное действие). Лишь незначительное число генов связано с проявлением только одного конкретного признака. У большинства высших животных и человека признаки определяются одновременным участием нескольких генов (полимерия). В той же мере отдельный ген может участвовать в развитии нескольких, казалось бы далеких признаков (плейотропия ). Последнее объясняет, почему при наследственных заболеваниях, возникающих в результате повреждения лишь одного гена, наблюдаются нарушения со стороны различных органов и процессов.
Для понимания закономерностей наследования признаков — порядка их распределения и комбинации в потомстве большое значение имело выяснение еще одного свойства генов — их способности к рекомбинации (см. Ген, обмен генами), т. е. перемещению из одной гомологичной хромосомы в другую. Это явление, изученное впервые Т. Морганом, обусловлено тем, что гомологичные хромосомы в созревающих половых клетках тесно сближаются (конъюгируют) и перекрещиваются. После этого хромосомы разъединяются, отдаляются друг от друга, а затем в процессе деления — мейоза (см. Размножение) — расходятся по разным клеткам. Места перекреста конъюгирующих хромосом (хиазмы) отчетливо видны под микроскопом. Т о., каждая хромосома после конъюгации и перекрещивания оказывается частично обновленной; она несет в себе часть наследственного материала другой хромосомы. В результате различные зрелые половые клетки оказываются разнокачественными в генетическом отношении. В этом сущность процесса перекомбинирования генетического материала. Процесс этот является одним из важных источников разнообразия организмов при скрещивании, т. к. рекомбинация генов влечет за собой различную комбинацию признаков у потомков (комбинативная изменчивость — см. Изменчивость).
Изучение сцепления и рекомбинации еще в те времена, когда не было данных о тонкой структуре хромосом и молекулярной структуре генов, позволило составить хромосомные карты, определить место гена в хромосоме, рассчитать расстояние между ними (так наз. составление генетических карт), предсказать результаты сочетания генов
а значит, и признаков при скрещивании.
Большое значение для проявления активности генов имеют условия существования данного индивидуума. Факторы окружающей среды оказывают большое влияние на реализацию генетической программы особи. Строение и функционирование любого организма определяется не только его наследственными потенциями, но и условиями, в к-рых эти потенции развиваются и реализуются.
Понимание закономерностей наследования было углублено при переходе к исследованиям на молекулярном уровне. Было выяснено, что материальной основой наследственности являются молекулы дезоксирибонуклеиновой к-ты (ДНК), а в молекулах рибонуклеиновых к-т (РНК) закодирована генетическая программа лишь у нек-рых вирусов (см. Нуклеиновые кислоты).
Молекулы ДНК входят в состав хромосом, они также имеются в ряде внутриклеточных органелл, находящихся в цитоплазме (митохондриях, пластидах растительных клеток и т. д.). В соответствии с этим говорят о ядерной и цитоплазматической (или внеядерной) Н.
Для реализации генетической программы организмы обладают способностью считывать генетическую информацию, закодированную в ДНК (см. Генетический код), в виде молекул информационных РНК (иРНК), являющихся копиями генов. В свою очередь молекулы иРНК, соединяясь со специальными структурами клеток — рибосомами (см. Клетка), программируют синтез белков, а последние управляют всеми реакциями в клетках, обеспечивая жизнедеятельность организмов.
Типы наследования. Построение генетических карт позволило выяснить характер и порядок расположения многих генов в хромосомах человека, а изучение проявления у организмов доминантных и рецессивных генов дало возможность вычленить несколько типов наследования. Прежде всего были изучены типы наследования признаков, кодируемых генами, расположенными в половых хромосомах. Важнейшей особенностью этого типа является то, что у гетерозиготного пола, несущего в каждой клетке тела две разные половые хромосомы — X и Y (напр., мужские особи у человека), гены, расположенные в одной из хромосом и не имеющие партнера в другой хромосоме, будут проявляться независимо от того, доминантны они или рецессивны. Поэтому, если у матери действие болезнетворного гена, расположенного в одной из Х-хромосом, может быть подавлено доминантным геном другой Х-хромосомы, то у ее сына, Получившего от нее Х-хромосому с болезнетворным геном, пусть даже и рецессивным, этот ген тем не менее может проявиться в виде наследственной болезни. Примером такой болезни является гемофилия (пониженная свертываемость крови и обусловленная этим кровоточивость), при к-рой мальчики, получившие от матери Х-хромосому с рецессивным болезнетворным геном, заболевают, а у девочек, получивших Х-хромосому с дефектным рецессивным геном от матери и нормальную Х-хромосому от отца, болезнь проявляться не будет. По этому же так наз. Х-хромосомному типу наследуются многие другие болезни — цветовая слепота, нек-рые формы диабета и др. Точно так же признаки, кодируемые генами, расположенными в Y-хромосоме и не имеющими аналогов в X-хромосоме, будут проявляться только у мужских особей, не передаваясь женским.
Иначе будет происходить наследование тех признаков, гены к-рых расположены не в половых хромосомах, а в аутосомах. В случае, если признак определяется доминантным геном, то любая хромосома матери или отца, попавшая при оплодотворении в зародышевую клетку, будет размножена во всех клетках тела развивающегося организма и даст проявление. Если речь идет о болезнетворном (патологич.) гене, то определяемый им признак проявится у любой из несущих его особей, как женской, так и мужской. В этом случае больные мальчики и девочки рождаются с одинаковой частотой. В случае брака между супругами, один из к-рых несет болезнетворный (патологич.) аутосомно-доминантный ген, вероятность проявления заболевания у их детей составляет 50%.
Гораздо более низка вероятность наследования признаков, кодируемых рецессивными генами, располагающимися в аутосомах. Такие гены широко распространяются в популяциях с помощью открытого советским генетиком С. С. Четвериковым процесса, позже названного генетическим дрейфом. В скрытом, рецессивном, состоянии действие этих генов подавляется их доминантными партнерами, лежащими в других парных хромосомах. Однако в случае, если каждый из родителей несет по одному одинаковому рецессивному гену, у 25% детей в каждой из парных хромосом будет иметься по паре рецессивных генов, и действие рецессивного гена проявится. Отсюда ясно, насколько вредно вступление в брак лиц, являющихся носителями одинаковых рецессивных генов. Хотя выявление такого носительства пока достаточно трудная задача, тем не менее очевидно, что наибольшая вероятность совпадения генотипов родителей будет наблюдаться при браках между близкими родственниками. По аутосомно-рецессивному типу наследуются многие тяжелые нарушения обмена веществ, заболевания нервной системы, болезни крови и др.
Проявление наследственных признаков организма может происходить на протяжении всего периода его индивидуального развития. Известно большое число наследственных болезни, проявляющихся не в раннем возрасте, а на более поздних стадиях развития. Так, напр., ряд наследственных форм диабета, тяжелое заболевание нервной системы — хорея Гентингтона и многие другие болезни проявляются в возрасте 30—40 лет, т. е. в то время, когда носители болезнетворных генов, не подозревая о своем дефекте, вступают в брак и дают потомство, также отягощенное этим дефектом. Отсюда ясна роль развития исследований по раннему выявлению носительства патологич. генов.
Важное значение имеет Н., обусловленная действием внехромосомного генетического материала, к-рый содержится в органеллах цитоплазмы (напр., в митохондриях). Совокупность цитоплазматич. генов (плазмогенов) обозначают термином «плазмон» (в отличие от совокупности хромосомных генов — генома). Плазмогены передаются потомкам гл. обр. по материнской линии, т. к. сперматозоиды, содержащие отцовские факторы, имеют ничтожно малое количество цитоплазмы, а следовательно, и малое количество плазмогенов. Внеядерные гены размножаются чаще всего независимо от хромосомного аппарата и потому их распространение подчиняется другим закономерностям. Изучению внеядерных генов уделяется особенно большое внимание, т. к. установлено, что многие важные признаки (повышенная способность к мутациям, устойчивость к антибиотикам и др.) определяются у микроорганизмов наследственными структурами цитоплазмы. Поскольку многие из этих микроорганизмов имеются и в теле человека (и часть из них может быть патогенной), то несомненно, что устойчивость к определенным антибиотикам будет приводить к устранению леч. эффекта антибиотиков и многим другим нежелательным последствиям. Установлено, что у высших организмов в каждой клетке значительная часть генов сосредоточена в ДНК митохондрий, с к-рыми связаны энергетич. процессы клеток, а также в других органеллах клетки, содержащих ДНК (напр., пластидах у растений). Естественно, что проявление активности этих цитоплазматич. генов играет важнейшую роль в жизнедеятельности организмов. Доказано также, что цитоплазматич. гены «работают» во взаимодействии с хромосомными и, т. о., взаимодействие генома и плазмона создает сложное и координированное во времени и пространстве проявление реакций и процессов, определяющих жизнедеятельность живых организмов.
Процессы реализации наследственной информации протекают в условиях постоянного воздействия на организм внешних по отношению к нему факторов среды. Для плода человека это прежде всего факторы материнского организма, внутри к-рого он развивается, и, конечно, факторы среды, действующие на организм матери, а через него и на плод.
Всякое нарушение процессов реализации генетической программы (генотипа) в совокупность признаков и свойств организма (фенотип ) может привести к необратимым нарушениям — болезням, порокам развития и др.
Жесткое детерминирование признаков генами проявляется далеко не во всех случаях, и потому генетики говорят о том, что гены, как правило, определяют не сами признаки, а норму реакции организма на изменяющиеся факторы внешней среды. В то же время изменения фенотипа, возникающие под действием окружающей среды и не закрепленные в генотипе, не наследуются потомками.
НАСЛЕДСТВЕННОСТЬ // Большая российская энциклопедия. Электронная версия (2017) // Новая версия
НАСЛЕ́ДСТВЕННОСТЬ, свойство организмов обеспечивать материальную и функциональную преемственность между поколениями. Н. реализуется в процессе наследования или воспроизведения в ряду поколений специфич. характера обмена веществ и индивидуального развития в определённых условиях внешней среды. Проявление Н. осуществляется в непрерывности живой материи при смене поколений. Поскольку организм развивается в результате взаимодействия генетич. факторов и условий существования, Н. может реализоваться в разл. вариантах в зависимости от особенностей генотипа и внешних условий. Напр., у особей с разным генотипом Н. может выражаться в одинаковом фенотипе, у организмов с одинаковым генотипом – в разных фенотипах (см. Модификации). Представление о Н. исторически возникло и развивалось как отражение существования материальной субстанции, обеспечивающей сходство организмов в ряду поколений. В связи с этим в генетической лит-ре появился ряд терминов, связывающих Н. с определёнными структурами клетки и объединяемых общим термином «генетический материал». После доказательства роли ядра в передаче признаков была сформулирована ядерная теория Н. В дальнейшем была разработана хромосомная теория Н., доказывающая, что наследств. факторы локализованы в хромосомах. По мере развития генетики выяснилось, что генетич. факторы могут находиться не только в ядре (хромосомах), но и в цитоплазме (некоторые органеллы клетки, плазмиды). В связи с этим возникло представление о цитоплазматической Н. Было также установлено, что генетич. информация хранится, воспроизводится и передаётся при размножении организмов в виде молекул нуклеиновых кислот (ДНК, РНК), являющихся материальными носителями всех видов Н. Особую роль в Н. играют точность воспроизведения молекул нуклеиновых кислот в процессах репликации и транскрипции и высокая степень точности синтеза белков в ходе трансляции. Функциональная преемственность между поколениями может обеспечиваться не только спец. материальными структурами, но и передачей информации от одного поколения другому в процессе обучения. Основа такого вида преемственности – условно-рефлекторная деятельность высших организмов, особенно человека. Для обозначения этого свойства был предложен термин «сигнальная наследственность». В ходе возникновения и развития жизни на Земле Н. играла решающую роль, обеспечивая закрепление достигнутых эволюционных преобразований. Благодаря Н. стало возможным существование разнообразных групп организмов как относительно самостоятельных, целостных систем (популяции, виды), сохранение приспособленности к определённым условиям существования. Именно поэтому Н. является одним из гл. факторов эволюционного процесса. Представляя в определённом смысле консервативность живых систем, Н. выступает в неразрывной связи с изменчивостью, определяя её возможные границы либо в процессе существования отд. особей (модификационная, онтогенетич. изменчивость), либо группы организмов в ряду поколений (генотипич. изменчивость). Изучение закономерностей Н. имеет важное значение для практики с. х-ва и медицины. Часто смешивают понятия «Н.» (свойство организмов) и «наследование» (процесс передачи признаков), что недопустимо.
Лит.: Жимулев И. Ф. Общая и молекулярная генетика. 4-е изд. М., 2007; Инге-Вечтомов С. Г. Генетика с основами селекции. 2-е изд. СПб., 2010.
