Советский Энциклопедический Словарь. 1980
Большая российская энциклопедия. 2016
Популярная медицинская энциклопедия. 1981
|
Вирус — Википедия |
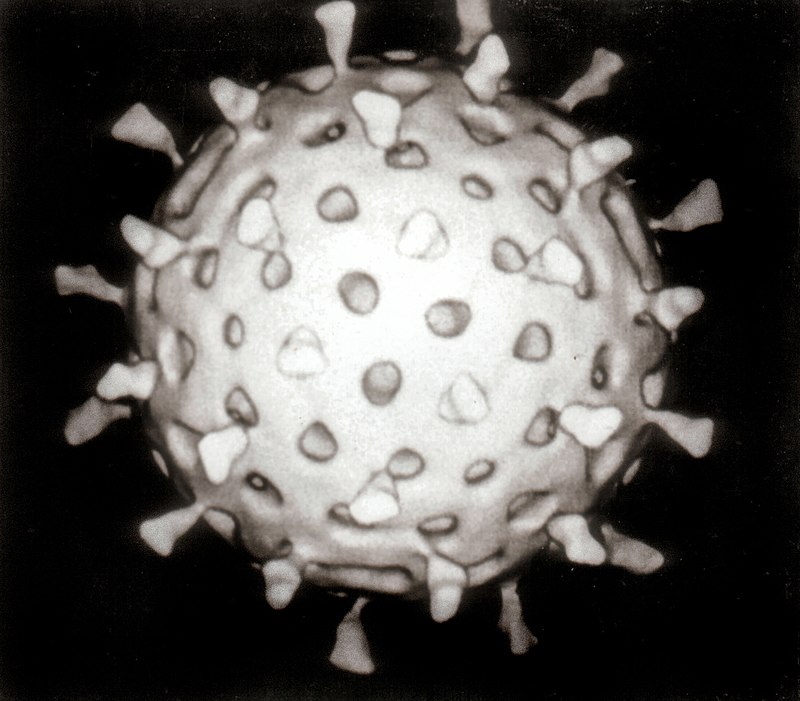
Энциклопедический Словарь. Том 1. — Москва, 1953—1955. — Стр. 309
ВИРУСЫ, возбудители инфекц. заболеваний, но размерам более мелкие, чем большинство известных микробов. Вследствие малых размеров В. проходят через бактериальные фильтры, отчего их называют фильтрующимися В. Значит. число инфекц. заболеваний у растений, животных и человека вызывается В. (мозаичная болезнь табака, чума рог. скота, ящур, грипп, оспа, корь и др.). Существование В. было впервые открыто русским учёным Д. И. Ивановским в 1892.
Советский Энциклопедический Словарь. — Москва, 1980. — Стр. 227-228
ВИРУСЫ (от лат. virus — яд), мельчайшие неклеточные частицы, состоящие из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки (капсида). Форма палочковидная, сферич. и др. Размер от 20 до 3000 нм и более. В.— внутриклеточные паразиты: размножаясь только в живых клетках, они используют их ферментативный аппарат и переключают клетку на синтез зрелых вирусных частиц — вирионов. Распространены повсеместно. Вызывают болезни р-ний, ж-ных и человека.
В. бактерий (бактериофаги) — классич. объект мол. биологии.
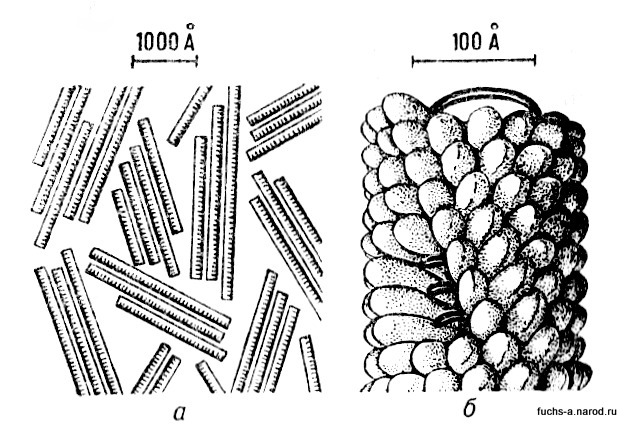
Вирус мозаичной болезни табака: а — электронная микрофотография; б — модель вирусной частицы.
ВИРУСОЛОГИИ ИНСТИТУТ им. Д. И. Ивановского АМН СССР, осн. в 194G (совр. назв. с 1950) в Москве. Иссл. по общей и мол. биологии вирусов, вирусным болезням.
ВИРУСОЛОГИЯ (от вирусы и ...логия), наука о вирусах. Общая В. изучает природу вирусов, их строение, размножение, биохимию, генетику. Мед., вет. и с.-х. В. исследует патогенные вирусы, их инф. свойства, разрабатывает меры предупреждения, диагностики и лечения вызываемых ими заболеваний. Раздел В., изучающий наследств. свойства вирусов, тесно связан с молекулярной генетикой.
Большая российская энциклопедия. Том 5. — Москва, 2006. — Стр. 374 // Электронная версия
ВИРУСЫ (от лат. virus – яд), мельчайшие биологич. объекты (обычно размером от сотых до десятых долей мкм). Важнейшая особенность В. – их неспособность к самостоятельному размножению, что связано с отсутствием у них механизмов для синтеза белков и производства энергии. Ранее считалось, что В. могут размножаться только в живых клетках (что обычно и имеет место), и это свойство входило в их науч. определение. Сейчас доказана принципиальная возможность размножения некоторых В. и в разрушенных клетках, но, несмотря на это, считается, что В. – облигатные (безусловные) внутриклеточные паразиты.
Строение вирусов
Вирусные частицы, или вирионы, могут иметь сферическую, нитевидную, палочковидную и более сложные формы. Одни В. (их иногда называют простыми) представлены только нуклеокапсидом – геномом В., заключённым в белковую оболочку (капсид), обычно построенную из множества молекул белка одного и того же или разных типов, уложенных по закономерностям кубической или спиральной симметрии. У других (сложных) В. нуклеокапсид окружён наружной оболочкой (мембраной), обычно состоящей из липидов с вкраплениями белков и гликопротеинов. Геном В. представлен либо молекулой ДНК (ДНК-содержащие В.), либо одной или несколькими молекулами РНК (РНК-содержащие В.), причём у разных В. эти молекулы могут быть однонитевыми или двунитевыми, линейными или кольцевыми. Размеры генома варьируют обычно от нескольких тысяч до 30 тыс. нуклеотидов у РНК-содержащих В. и до нескольких сотен тысяч нуклеотидов у ДНК-содержащих. Самые мелкие геномы имеют всего неск. генов, которые кодируют белок (белки) вирусной оболочки и некоторые ферменты, принимающие участие в синтезе вирусных нуклеиновых кислот. В более крупных геномах (особенно у ДНК-содержащих В.) закодирована дополнительная информация, не всегда обязательная для сохранения жизнеспособности В., но обеспечивающая их более эффективное и независимое от внутриклеточных условий размножение. Напр., в ДНК-геноме мимивируса, поражающего амёб, заключено ок. 1200 генов, т. е. больше, чем в ДНК некоторых паразитич. микроорганизмов. Существуют т. н. дефектные В. (напр., В. гепатита дельта), которых называют паразитами «вдвойне»; у них нарушены те или иные системы репродукции, поэтому для образования потомства им требуется не только внутриклеточная среда, но и помощь со стороны др. полноценного В. (в данном случае – В. гепатита B). Для некоторых парвовирусов в качестве помощника выступают аденовирусы. В. могут поражать все живые организмы. В. бактерий называют бактериофагами (фагами). Описано неск. тысяч В. Их относят к десяткам семейств, среди которых выделяют роды и виды.
Размножение вирусов
Размножение вирусов протекает в неск. стадий. Сначала В. прикрепляется к клетке, что обычно требует наличия химич. сродства между поверхностью вирусной частицы и тем или иным компонентом клеточной поверхности (рецептором). Это взаимодействие весьма специфично, что является одной из причин высокой избирательности взаимодействия В. с клеткой (напр., В. полиомиелита или В. иммунодефицита человека – ВИЧ в естеств. условиях инфекционны только для человека). Но есть В. и с широким спектром хозяев, заражающие, напр., млекопитающих и насекомых (так, В. клещевого энцефалита размножается в организме человека и клеща). В клетки растений В. может проникать только после механич. повреждения (напр., насекомыми) клеточной стенки, т. к. она служит для них непреодолимым барьером. После взаимодействия В. с рецептором его геном (в свободном виде или в комплексе с белками) проникает внутрь клетки, где начинается синтез вирусных нуклеиновых кислот и белков (структурных компонентов вириона, а также вирусных ферментов и др. молекул, способствующих эффективному размножению вируса благодаря их взаимодействию с вирусными или клеточными макромолекулами). Последовательность образования этих двух типов молекул у разных В. варьирует. Однако всегда синтез вирусных белков направляется вирусными нуклеиновыми кислотами и происходит на рибосомах клетки. Синтез же вирусных нуклеиновых кислот осуществляется при участии либо вирусных, либо клеточных ферментов, нередко и тех и других. Затем вирусные нуклеиновые кислоты и структурные белки объединяются с образованием дочерних вирионов, которые покидают клетку, разрушая её или сохраняя целостность. Клеточная плазматическая мембрана сохраняется, напр., если В. покидает клетку путём т. н. почкования: «незрелый» вирион сначала обволакивается клеточной мембраной, включающей некоторые из вирусных белков, затем «отшнуровывается», а целостность клеточной мембраны восстанавливается. Т. о., для построения оболочки В. могут использоваться клеточные липиды и углеводы. Инфекц. процесс, инициированный одним вирионом, может приводить к образованию до 103–105 дочерних вирусных частиц.
Выделяют неск. осн. типов взаимодействия В. и клетки. При продуктивной литической инфекции заражённая клетка после образования вирусного потомства гибнет. В случае хронической персистентной (стойкой) инфекции заражённая клетка продолжает жить и делиться, постоянно поддерживая размножение В., хотя её функции могут претерпевать некоторые изменения. Такая инфекция может продолжаться длительное время без заметных внешних проявлений, не вызывая признаков заболевания, хотя могут иметь место определённые изменения регуляции клеточных процессов. Особый тип взаимодействия с клеткой наблюдается у т. н. умеренных В., к числу которых относятся бактериофаг лямбда и ряд В. человека и животных (напр., ВИЧ). Их геном ковалентно встраивается (интегрируется) в клеточную хромосому, превращаясь в элемент клеточного генома, и теряет способность к автономной репликации. В таком состоянии В. называют провирусом, а бактериофаг – профагом. У умеренных РНК-содержащих вирусов (ретровирусы) геном предварительно переходит в ДНК-форму. Экспрессия вирусных генов происходит так же, как и у генов клетки, а кодируемые ими белки могут влиять на свойства клетки, напр. на регуляцию её деления. В некоторых случаях встраивание вирусного генома в геном клетки может приводить к почти полному угнетению экспрессии клеточных генов. При этом клетка сохраняет жизнеспособность, но её свойства могут существенно меняться (напр., может произойти её злокачественная трансформация). Патологич. изменения клетки могут быть также связаны с угнетением синтеза РНК и клеточных белков, нарушением мембранной проницаемости и др. процессами, заканчивающимися во мн. случаях смертью клетки (цитопатический эффект).
В клетке и в целом организме существуют механизмы, препятствующие размножению В. и развитию вирус-индуцированной патологии, в т. ч. врождённый (напр., индукция интерферонов) и приобретённый (напр., образование антител) иммунитет. В свою очередь, мн. В. располагают механизмами, противодействующими клеточной защите. Так, некоторые В. подавляют секрецию интерферона заражённой клеткой, облегчая тем самым заражение соседних клеток вирусным потомством. Исход инфекции и судьба заражённой клетки и организма зависят от баланса противоположно направленных активностей В. и клетки. Поражение клеток в заражённом организме ведёт к нарушению работы соответствующих органов и тканей, проявляющихся в виде вирусных заболеваний. Некоторые из них имеют характер массовых эпидемий и даже пандемий и могут сопровождаться значительной смертностью (в т. ч. СПИД, грипп, ящур и др.). Важнейшим средством профилактики вирусных заболеваний является специфическая вакцинация.
Генетическая изменчивость вирусов, их эволюция
Изменчивость В. объясняет их способность «уходить» от иммунного ответа заражённого им организма (напр., в случае В. гриппа) и легко образовывать варианты, резистентные к лекарственным препаратам. Результатом изменчивости может быть также приобретение способности вызывать заболевания человека В., ранее патогенными только для животных. Нередко обнаруживается явное родство между отд. белками (напр., ферментами, осуществляющими синтез нуклеиновых кислот) у В., которые, на первый взгляд, не имеют между собой ничего общего. В ряде случаев заметно сходство между вирусными и клеточными белками. Из этого следует, что в ходе эволюции происходил (и, по-видимому, происходит) обмен генетич. информацией как между разными В., так и между В. и клеточными организмами. Ярким примером явления трансдукции – способности В. переносить гены или их фрагменты от одного организма к другому – может служить бактерия дифтерийная палочка, которая начинает вырабатывать токсин только после заражения её определённым фагом. Такой перенос генов (его называют горизонтальным) пытаются использовать для генной терапии путём создания на основе В. искусств. конструкций – векторов, способных вводить в заражаемую клетку здоровые или корректирующие гены.
Особую категорию составляют т. н. эндогенные В. Их геном в виде двунитевой ДНК, соответствующей геному ретровирусов, постоянно находится в составе клеточной хромосомы и функционирует как набор клеточных генов. Эндогенные В. могут быть полноценными, и тогда их экспрессия приводит к образованию В., способного заражать др. клетки (напр., В. рака молочных желёз мышей). В большинстве случаев, однако, эндогенные В. имеют дефектный геном, в котором некоторые вирусные гены отсутствуют или повреждены. Генетич. материал разнообразных эндогенных В. составляет весьма значительную долю хромосомной ДНК животных, в т. ч. человека. Они обнаружены и у растений. Считается, что эндогенные В. возникли в результате заражения половых клеток и последующей интеграции вирусной и клеточной ДНК. Такое заражение в процессе эволюции могло происходить многократно, и многие эндогенные В. считаются весьма древними.
Ввиду огромного разнообразия В. построение их эволюционного «древа» весьма затруднено. Среди мн. предположений о происхождении В. наибольшего внимания заслуживают два: или они возникли из обособившихся («одичавших») элементов клеточного генома, или в какой-то форме существовали уже на доклеточной стадии биологич. эволюции.
В. сыграли и продолжают играть выдающуюся роль как удобные модельные объекты для изучения общих закономерностей молекулярной биологии. Именно при изучении В. были расшифрованы важнейшие закономерности синтеза белков и нуклеиновых кислот и регуляции этих процессов, сформулированы мн. понятия молекулярной биологии и молекулярной генетики. См. также ст. Вирусология и лит. при ней.
Популярная медицинская энциклопедия. — М.: Советская энциклопедия, 1981
ВИРУСЫ — мельчайшие возбудители инфекционных заболеваний человека, животных, растений и бактерии, являющиеся внутриклеточными паразитами, не способными к жизнедеятельности вне живых клеток. Вирусные инф. болезни широко распространены. Еще Демокрит (4б0 — ок. 370 г. до н. э.) и Аристотель (384—322 г. до н. э.) описали клинич. картину бешенства. С древних времен известны оспа, полиомиелит, грипп. Однако только в 1892 г. рус. ученый Д. И. Ивановский обнаружил болезнетворное начало, проникавшее сквозь фильтры, задерживающие остальные виды микроорганизмов.
Долгое время дискутировался вопрос, являются ли В. живыми организмами или представляют собой только комбинацию гигантских молекул. В настоящее время выяснено, что В. являются наиболее примитивными существами, но обладают всеми основными свойствами живых организмов. Они размножаются, т. е. воспроизводят потомство, сходное с исходными родительскими В., хотя способ их размножения весьма своеобразен и во многом отличается от того, что нам известно о размножении других живых существ. Обмен веществ у В. тесно связан с обменом веществ зараженных клеток. Они обладают наследственностью, свойственной всем живым организмам, к-рая обусловлена теми же биол. и хим. структурами, что и у других живых организмов, т. е. нуклеиновыми кислотами. Наконец, В., как и все другие живые существа, обладают изменчивостью и приспособляемостью к меняющимся условиям окружающей среды.
В. могут существовать в двух формах: внеклеточной (покоящейся) и внутриклеточной (размножающейся). Существующие вне клеток вирионы (вирусные частицы) не обнаруживают никаких признаков жизни. Попав в организм, вирионы проникают в чувствительные к ним клетки и переходят из покоящейся в размножающуюся форму. Начинается сложное и многообразное взаимодействие В. и клетки, заканчивающееся образованием и выходом в окружающую среду дочерних вирионов.
Строение и химический состав вирионов. Долгое время о существовании В. судили по их болезнетворному действия. Непосредственно наблюдать В. можно только с помощью электронного микроскопа, дающего увеличение в десятки и сотни тысяч раз (максимальное увеличение обычного оптич. микроскопа — 1500 раз). Впервые увидеть В. удалось лишь через 50 лет после того, как они были открыты.
Самые крупные — В. оспы достигают величины 400—700 нм (нм — миллионная доля мм) и приближаются по размерам к небольшим бактериям самые мелкие (возбудители энцефалита, полиомиелита, ящура) измеряются всего десятками нм, т.е. близки к крупным белковым молекулам (напр., молекуле гемоглобина крови), т. о., среди В. есть свои великаны и карлики. По внешнему виду тельца В. (вирионы ) напоминают кубики, палочки, шарики и нити. В., поражающие бактерии (см. Бактериофаг ), похожи на головастиков и состоят из головки и хвоста (рис. 1).
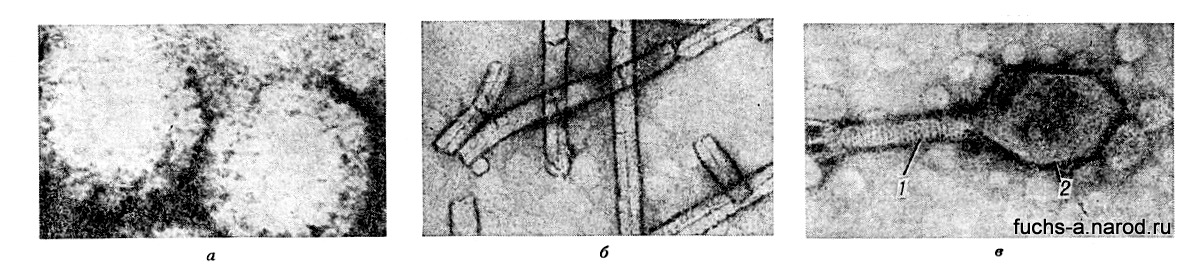
Рис. 1. Различные формы вирионов (электронно-микроскопическая картина, х 3000 000): а — шаровидные реовирусы человека: б — палочковидный вирус табачной мозайки; в — бактериофаг, состоящий из хвоста (1) и головки (2).
Самые простые вирусы состоят из белков и нуклеиновой к-ты. Наиболее важная часть вирусной частицы — нуклеиновая к-та, носитель генетической информации. Клетки человека, животных, растений и бактерий всегда содержат два типа нуклеиновых к-т дезоксирибонуклеиновую (ДНК) и рибонуклеиновую (PHK) (см. Нуклеиновые кислоты), В.— лишь один тип — или ДНК, или РНК. Так, В. оспы, аденовирусы и большинство бактериофагов содержат ДНК, а В. гриппа, полиомиелита, энцефаломиелитов — РНК, к-рая у В. (в отличие от РНК клеток животных) содержит генетическую информацию. Имеются также В., содержащие однонитчатую ДНК (нек-рые бактериофаги) и двунитчатую РНК (реовирусы), к-рые не встречаются среди клеточных нуклеиновых к-т. Второй обязательный компонент вирусной частицы — белки, сложность строения к-рых у разных В. различна.
Более сложные по структуре В., кроме белков и нуклеиновых к-т, содержат углеводы, липиды. Для каждой группы В. характерен свой набор белков, жиров, углеводов и нуклеиновых к-т. Многие В. содержат также ферменты.
Каждый компонент вирусной частицы имеет определенные, весьма сложные функции: нуклеиновая к-та определяет наследственные и инф. свойства и играет ведущую роль в изменчивости В., а ферменты участвуют в их размножении. Обычно нуклеиновая к-та находится в центре частицы и защищена от неблагоприятных воздействий белковой оболочкой, как бы одета в нее. Оболочка (капсид) состоит из определенным образом уложенных однотипных белковых молекул (капсомеров ), к-рые образуют симметричные геометрические формы. В случае кубической симметрии нить нуклеиновой к-ты свернута в клубок, а белковые капсомеры плотно уложены вокруг нее. Такие вирусные частицы напоминают малину и по форме приближаются к шарику. Так устроены В. полиомиелита, ящура, адено-, рео-, риновирусы и др. При спиральной симметрии нить нуклеиновой к-ты В. закручена в виде спирали, каждый виток к-рой покрыт капсомерами, тесно прилегающими друг к другу (нуклеокапсид). Вирусные частицы в этом случае напоминают початок кукурузы и по форме приближаются к палочке. Типичный пример — вирус табачной мозаики.
У сложно устроенных В. сердцевина в виде туго свернутой спирали покрыта одной или несколькими внешними оболочками, в состав к-рых входят различные вещества. Такое строение имеют, напр., орто- и парамиксовирусы. Сложнее устроен В. оспы (рис. 2).
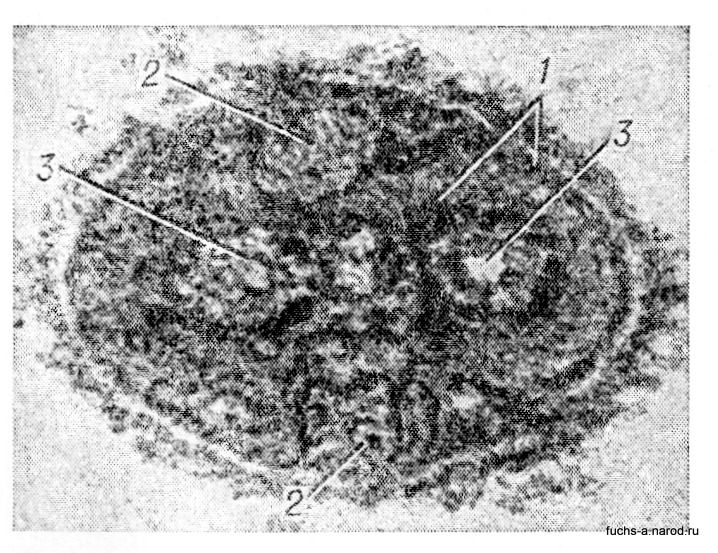
Рис. 2. Сложно устроенный вирион оспы (электронно-микроскопическая картина, х 200 000): 1 — внешняя и внутренняя оболочки; 2 — боковые тела; 3 — сердцевины вириона.
В его центре находится «внутреннее тельце», напоминающее ядро клеток высших организмов. Опо состоит из белка и ДНК и окружено белковым слоем, по бокам от к-рого имеются два образования также белковой природы. Снаружи вирусная частица покрыта оболочкой. Довольно сложно устроены и частицы В. гриппа. Нуклеокапсид в виде трубок уложен внутри частиц и одет в оболочку. Особенно подробно изучено строение В. бактерий — бактериофагов, к-рые состоят из головки и хвоста. Различные типы фагов отличаются друг от друга размерами, а также формой головки, к-рая может быть палочковидной,круглой,овальной,в виде пяти- или шестиугольника. Хвост фага одет белковым чехольчиком, от к-рого отходят длинные тонкие волокна, играющие роль присосок при прикреплении частицы фага к бактерии.
Размножение вирусов. Взаимодействие вириона с живой клеткой осуществляется в несколько этапов. В начальный (подготовительный) период вирион прикрепляется к клетке, проникает внутрь ее, после чего белковая оболочка вириона разрушается, освобождая нуклеиновую к-ту (рис. 3).
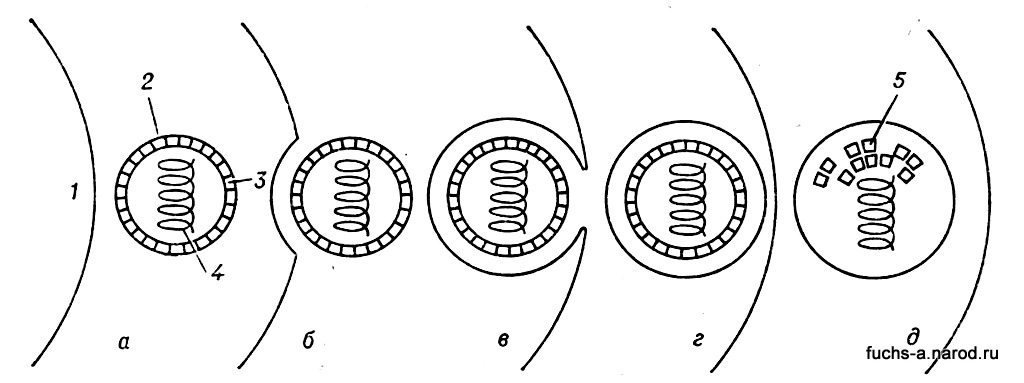
Рис. 3. Схема начального этапа взаимодействия живой клетки (1) с вирионом (2): а — клетка и вирион (3 — оболочка вириона, 4 — вирусная нуклеиновая кислота); б — начальная стадия адсорбции вириона на клетке (видно небольшое впячивание на поверхности клетки); в — вдавленная внутрь клеточная оболочка окружает вирион; г — вокруг вириона образовалась замкнутая вакуоль; д — оболочка вириона разрушена (видны ее фрагменты — 5), освободилась вирусная нуклеиновая кислота.
Наступает скрытый (латентный) период вирусной инфекции, во время к-рого присутствие в зараженной клетке вирусных частиц нельзя обнаружить никакими методами — родительский вирион как бы исчезает. В этот период проникшая в клетку вирусная нуклеиновая к-та организует синтез вирусных компонентов потомства, используя для этой цели ферментативную систему хозяина. Цикл размножения заканчивается формированием дочерних вирионов и выходом их из клетки (конечный период).
Атаки на клетку начинаются с прикрепления В. к клеточной стенке, или так наз. адсорбции вируса. Она отличается большой специфичностью: каждый В. способен прикрепляться лишь к определенным клеткам, имеющим на своей поверхности спец. рецепторы. На одной клетке могут адсорбироваться десятки и даже сотни вирусных частиц. Затем начинается внедрение вирионов, осуществляемое самой клеткой (виропексис). Она как бы «втягивает» прикрепленные к ее оболочке вирусные частицы внутрь. Здесь мы встречаемся с тем случаем, когда созданное многовековой эволюцией целесообразное приспособление приводит к совершенно нежелательным результатам. Действительно, активный захват клеткой из окружающей среды различных частиц (фагоцитоз) или капелек воды (пиноцитоз) имеет большое значение для ее нормальной жизнедеятельности, а использование этих же механизмов для захвата В. скорее напоминает самоубийство. Если бы дело ограничилось только простым «проглатыванием» вирусной частицы, это не приводило бы к трагическому для клетки финалу. Но механизм фагоцитоза предусматривает также последующее растворение фагоцитированной частицы (в нашем случае В.). Для этой цели клетка использует имеющийся у нее набор спец. ферментов, разрушающих вирусные белки. Освобожденные нуклеиновые к-ты В. по клеточным каналам очень быстро проникают в ядро клетки или остаются в цитоплазме.
Более просто устроенные бактерии не способны сами захватывать частицы из окружающей среды. Поэтому у бактериофагов имеются спец. приспособления для преодоления плотной бактериальной стенки. В концевой части хвоста содержится особый фермент, к-рый растворяет бактериальную оболочку. Затем микроскопические «мышцы» хвоста сокращаются и нуклеиновая к-та фага «впрыскивается» внутрь клетки, происходит как бы инъекция с помощью шприца. В результате белковый чехол фага остается на поверхности клетки, а внутрь клетки попадает лишь нуклеиновая к-та.
Нуклеиновые к-ты В. осуществляют программу по созданию в клетке нового вирусного потомства. Это было доказано оригинальными опытами. Удалось разделить В. на составляющие их компоненты — белки и нуклеиновые к-ты. Оказалось, что заражение клеток и размножение В. происходило только после добавления к клеткам вирусной нуклеиновой к-ты. Иными словами, нуклеиновые к-ты В. сами по себе могут вызывать размножение В., т. е. обладают инфекционными свойствами. В другом опыте два В. были разделены на составляющие компоненты, а затем «переодетый» нуклеиновую к-ту одного В. «одели» в оболочку другого. Полученными гибридами были заражены чувствительные клетки. Было обнаружено, что оба «переодетых» В. способны размножаться, а образующееся потомство всегда подобно тому В., нуклеиновую к-ту к-рого содержал гибрид.
Проникшая в клетку вирусная нуклеиновая к-та управляет всеми процессами размножения В. Сначала она заставляет клетку синтезировать так называемые ранние белки, подавляющие собственный обмен веществ клетки и обеспечивающие синтез нуклеиновых к-т дочерних частиц. Образование их происходит в результате самокопирования родительской нуклеиновой к-ты. Генетическая информация, заложенная в нуклеиновой к-те В., определяет состав белков, из к-рых строятся дочерние частицы так наз. поздних белков. В ДНК-содержащих В. реализация этой информации осуществляется обычным для клетки путем: на ДНК синтезируется информационная РНК (транскрипция), управляющая последующим биосинтезом белков (трансляция). В нуклеиновой к-те многих РНК-содержащих В. объединены и генетическая, и информационная функции: РНК участвует и в репликации, и в трансляции (в воспроизводстве нуклеиновых кислот и белка В.).
У многих В. построение белковых оболочек и внутреннего содержимого идет раздельно. Клетка «зарабатывает» отдельные детали, к-рые потом соединяются, образуя вирусные частицы. Когда в зараженной клетке накопится достаточное количество «заготовок» для будущих вирусных частиц, наступает как бы сборка деталей (композиция). Процесс этот происходит обычно вблизи клеточной оболочки, к-рая принимает в нем участие (рис. 4).
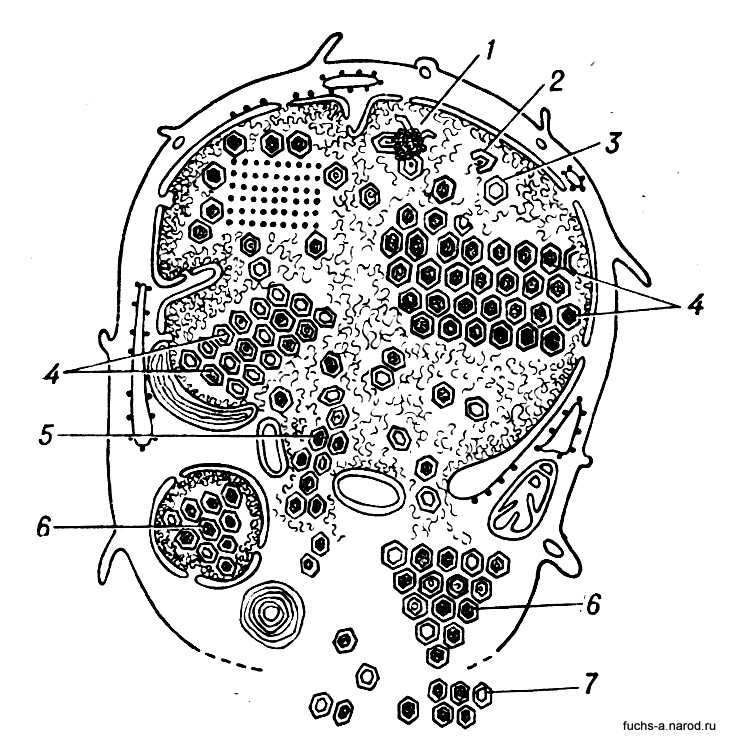
Рис. 4. Схема размножения аденовирусов в живой клетке: 1, 2, 3 — формирование дочерних вирионов в ядре клетки; 4 — внутриядерное скопление вирионов в виде кристалов; 5 — переход дочерних вирионов из я
